|
||||
|
|
||||
|
Estado del metabolismo de los lípidos en la infección por VIH-1 y SIDA Dres.
Raúl Gutierrez-Rodriguez, MD (*) 1. Estado de los Lípidos en relación a la Progresión de Enfermedad por VIH-1 1.1 Perfil Lipídico en la Infección por VIH-1 y SIDA Conforme la infección por VIH-1 avanza, ella se caracteriza por anormalidades metabólicas y nutricionales que incluyen hipertrigliceridemia, hipocolesterolemia, pérdida de peso, hipermetabolismo y pérdida de masa corporal magraT. El modelo de alteraciones en los niveles séricos de lípidos es semejante al reportado para otras infecciones("". Estas alteraciones se encuentran asociadas a procesos ininunológicos como son la proliferación y diferenciación de células linfoides, activación monocitaria inducida por endotoxina, liberación de citoquinas, y maduración de linfocitos T hasta convertirse en células supresoras y en células asesinas ("'). La hipertrigliceridemia, y la hipocolesterolemia han sido descritas como las alteraciones más comunes en el metabolismo de los lípidos durante la infección por V11-1- 1 y SIDÑ', "'). Otros estudios han demostrado niveles reducidos de lipoproteínas de baja y alta densidad (LDL y HDL, respectivamente), lípidos totales y ácidos grasos, además de niveles altos de metabolitos del ácido araquidónico(22:4-n-6y22:5-Q-6)("'). Asímismo, se han reportado niveles elevados de ácidos grasos polinsaturados (PUFAs) en pacientes con conteo celular CD4' por encima de 400/nim~ así como hipertrigliceridemia observada en estadíos avanzados de la infección por VIH-1 (con conteo celular CD4 por debajo de 50/mm')('), lo que sugeriría que conforme la enfermedad avanza, se encontraría asociada a niveles sanguíneos alterados de triglicéridosy ácidos grasos polin saturados (". Las alteraciones en el perfil lipídico plasmático en sujetos infectados con VIH-1 están asociadas a progresión de la enfermedad y reflejan claramente la influencia de la homeostasis lipídica sobre la función inmunológica"`. Edginton et al, observaron propiedades bioreguladoras de las lipoproteínas plasmáticas sugiriendo que, tanto el colesterol - HDL como los niveles altos de triglicéridos, afectan la inmunocompetencia de los sistemas humoral y celular (4) . 1.2 Hipertrigliceridemia, Progresión de Enfermedad por VIH-1 y Mortalidad La hiperlipidemia ha sido un hallazgo común en varias infecciones bacterianas y parasitarias`. En algunas infecciones bacterianas los niveles de triglicéridos se encuentran elevados durante la fase febril aguda`, mientras que en otras infecciones causadas por virus, los niveles de triglicéridos se incrementan durante la fase inmediata de convalecencia"'. Se ha reportado niveles bajos de LDL y HDL en la fase aguda de la enfermedad, previo a la detección de niveles elevados de triglicéridos". Se han observado niveles persistentemente bajos de colesterol total durante la fase de convalecencia de una infección, cuando los niveles de triglicéridos se incrementan. En tanto que en infecciones vírales experimentales, se ha observado un descenso en los niveles de colesterol antes de la elevación de los niveles de triglicéridos ". Durante los estadíos tempranos de la enfermedad por VIH-1, los triglicéridos parecen no registrar alteraciones en sus niveles plasmáticos"`. Nuestras investigaciones llevadas a cabo en 94 hombres homosexuales infectados con el VIII- l(7,14)9 han demostrado que los niveles promedio de triglicéridos (100.1±50.8 mg/dl) se encontraban dentrodel rango considerado como normal «l50 mg/dl) en el 84% de los hombres VIH- 1 positivos, y que los niveles de triglicéridos no registran diferencia significativa respecto a los controles seronegativos para el VIH- 1 (111.9 ± 62.9 mg/dl). Además, el análisis de regresión longitudinal de los datos, recolectados a lo largo de 18 meses en esta cohorte, mostró que los niveles de triglicéridos no estuvieron asociados a otros parámetros considerados predictores de progresión de enfermedad hacia el SIDA tales como, descenso en los niveles de CD4'y B,-microglobulina, ni tampoco relacionados con el estadío de la enfermedad "'. Así, los niveles de triglicéridos parecen estar dentro del rango normal durante los estadíos tempranos de la enfermedad por VIH-1, con una variación hacia el incremento de los niveles conforme la infección avanza, resultando finalmente en hipertriglíceridenúa ('). En el SIDA, los niveles plasmáticos de triglicéridos se encuentran notablemente elevados"'). Se especula que la acumulación de triglicéridos podría ser una consecuencia de los trastornos metabólicos producidos por algunas citoquinas [e.g. Factor de Necrosis Tumoral (TNF- a)] liberadas durante la activación del sistema inmunológico(2"'). Algunos de estos trastornos han sido. relacionados con alteraciones hepáticas y del mecanismo periférico de depuracióé). Estos mecanismos incluyen: 1. Incremento de la síntesis de novo de ácidos grasos lo que conduce a un incremento en la secreción de lipoproteínas de muy baja densidad (VLDL), 2. Incremento de la lipólísis en el tejido adiposo, reesterificando los movilizados ácidos grasos hasta triglicéridos en el hígado los que, en vez de ser oxidados, son secretados nuevamente como VLDL, 3. Niveles disminuidos de lipoproteinlipasa (LPL) originando una depuración disminuída de liproteínas ricas entriglicéridos""). Grunfeld y Koder ">, observaron que la hipertrigliceridemia constituye un hallazgo común en un amplio espectro de condiciones clínicas entre pacientes con SIDA, y sugirieron que la infección VIH-1 por sí misma, además de infecciones oportunistas específicas, podría ser un estímulo para (14,15) producir un estado de hipertrigliceridemia. Estudios in vitro han demostrado que niveles plasmáticos elevados de triglicéridos favorecen el crecimiento de especies tipo mycobacteria, y en el caso del SIDA, se ha postulado que la hipertrigliceridemia podría contribuir a la replicación descontrolada del Mycobacterium avium intracellulare (MAI)("). Así mismo, la naturaleza progresiva de la hipertrigliceridemia ha sido identificada como un parárnetro sensible de predicción de mortalidad en estudios relacionados al (17,18) SIDA en pacientes adultos y pediátricos González - Clemente, sugiere que la asociación entre hipertrigliceridemia y mortalidad podría estar explicada por la relación entre los niveles altos circulantes de triglicéridos, los de interferon alfa (IF-u), y los de antígeno p24 del VIH- 1 ("~. Otros mecanismos de acción, propuestos para explicar la relación entre los perfiles lipídicos, progresión de enfermedad por VIH y mortalidad, incluyen la habilidad de los lípidos plasmáticos para influenciar la función celular a través de cambios en (4) la composición lipídica de la membrana celular . Además, se estitna que los cambios en la membrana celular afectan la replicación (20,21) y la activacion inmunológica del VIH- 1 ('). Se requiere de más estudios de investigacion para determinar la relación que hay entre los disturbios del metabolismo lipídico, las alteraciones hormonales, la influencia de citoquinas, y la forma en que estos cambios afectan el desarrollo de la malnutrición y alteraciones del sistema inmunológico", 1.3 Hipocolesterolemía y Enfermedad por VIH-1. Algunos estudios han documentado que en los primeros estadíos de la infección por VIH-1, existe disminución en los niveles de colesterol total (CT), así como del contenido en las lípoproteínas de alta y baja densidad (HDL y LDL, respectivamente)","). Grunfeld et al, han reportado niveles bajos de CTHDL (incluyendo HDL, y HDL,) y LDL en individuos infectados con VIH- 1 y en pacientes con SIDA ("'. Los transtomos del metabolismo del colesterol, preceden al desarrollo de hipertrigliceridetnia durante la infección por VIHl("). En un estudio transversal conducido en 63 individuos infectados con VIVI- 1, Zangerle et al (') hallaron niveles de colesterol-LDL por debajo del rango normal en el 55 % de casos, y niveles bajos de colesterol-HDL en el 46% de los casos estudiados. El mismo estudio reportó niveles disminuidos de colesterol LDL y HDL en todas las etapas de la infección por VIH-1, pero con más frecuencia en individuos con niveles bajos de conteo celular CD4' (8) Nuestros estudios (7,14) también han demostrado significativamente niveles bajos de CT, HDL y LDL en una población homosexual masculina infectada con VIH-1 al ser comparada con un grupo control no infectado (p<0.05). En el 40% del grupo infectado se pudo observar hipocolesterolemia « 150 mg/dL) asociada a su vez 18,25) con niveles elevados de 82-inicroglobulina (7,18,25). Conforme los niveles de B,-microglobulina tienden a estar asociados con disminución futura del conteo celular CD4"), nuestros resultados sugieren que las determinaciones séricas de colesterol podrían servir como un índice de severidad de la infección por VIH- 1 ('). La ocurrencia simultánea de niveles bajos de colesterol con niveles altos de triglicéridos podría afectar la imnunocompetencia de los sistemas humoral y celula?). Los cambios en la membrana celular podrían producirse también por a hipocolesterolemia la que a su vez podría afectar la replicación vital"". La disminución de niveles de colesterol-HDL observada en la infección por VIH1 podría contribuir a la presencia del sincitio (fusión celular) induciendo a su vez, la expresión de fenotipos del VIH-1"". Todos estos mecanismos pueden contribuir a las relaciones entre los niveles plasmáticos de lípidos con la activacion inmunológica y progresión de la enfermedad
2. Alteraciones del Metabolismo de los Upídos en la Infección porVIR-lySIDA 2.1 Acidos Grasos Los ácidos grasos polinsaturados (PUFAs) juegan un papel importante en la respuesta inmunológica mediante la regulación de los cambios en la membrana celular de células imnunocompetentes y síntesis de prostaglandinas y AMP (21) . Estudios in vitro han observado que los ácidos linolénico y araquidónico pueden suprimir la transformación linfocítica lectina - inducida, influenciando directamente en la producción de (1,21) prostaglandina E~ . En otros estudios, la prostaglandina E2 (cuyo precursor es el ácido linoleico) ha ejercido una acción supresora sobre la función celular T, en respuesta a antígenos y mitógenos, la generación de células citotóxicas y producción de linfoquinas(). Algunos PUFAs son efectivos en la destrucción de virus que poseen componente lipídicos en su envoltura. EL VIH- 1 pertenece a ésta clase de virus, aunque el efecto del PUFA sobre el virus aún no ha sido descrito(2'). Nuestro estudio conducido en 36 varones homosexuales infectados con VIH-1 y en 17 varones homosexuales no infectados, como grupo control, demostró que al medir la concentración total de ácidos grasos plasmáticos, el perfil de PUFAs en el grupo infectado era diferente, pero no significativamente alterado, del registrado en el grupo control durante los estadíos tempranos de la infección por VIH-1. La cantidad total promedio de 0-6-PUFA (18:2 y 20A) fue menor en el grupo infectado al compararse con el control, y el índice promedio de ácido linolénico /araquidónico (18:2 a 20A) fue más alto en el grupo infectado. Estos cambios fueron correlacionados con índices de función inmune. La respuesta a fitohemaglutinina (PHA), un agente mitogénico que induce blastogénesis linfucitaria, fue inversamente proporcional a la concentración plasmática total de 0-6-PUFA, y directamente proporcional al índice linolénico /araquidónico en el grupo infectado, sugiriendo que los niveles reducidos de ácido araquidónico podrían estar asociados a la preservación de la respuesta linfocitaria normal a la PHA"). Se ha descrito cambios en los fasfalípidos de la membrana celular linfocitaria en relación a la progresión de enfermedad por VIH-l("). En un estudio conducido en homosexuales asintomáticos infectados con VIH-1 se observó un incremento de los niveles de ácido linolénico. En contraste, los pacientes sintomáticos con conteo celular CD4' por debajo de 500xlO'/L, exhibieron una disminución en los niveles de ácidos linolénico y araquidónico junto con un incremento de los niveles de ácido oleico. Estos hallazgos sugieren que una disminución en las concentraciones de ácidos grasos fi-6 puede representar la activación de la vía de la cieloxigenasa; la vía metabólica implicada en la biosíntesis de prostaglandinas (6,22) . La citopatología del VIH- 1 puede depender no solo de la interacción de la glicoproteína de superficie gp 120 (uno de los principales componentes de la cubierta del virus) con el receptor CD4, sino también del incremento de la fluidez de la membrana celular debido a un aumento de la concentración de ácido oleico (29) . En un estudio conducido por Begin et a l(28), los niveles plasmáticos absolutos de los ácidos linoleico, oleico y esteárico, fueron significativamente más bajos en pacientes con SIDA (n=11) que en aquellos individuos no infectados con el VlH-l("), probablemente, debido al estadío de la enfeimedad(6,28). Estudios in vitro llevados a cabo en cultivos tisulares han sugerido que el VIH-1 altera el índice ácido esteárico/ácido oleico en las membranas celulares, produciéndose una marcada desaturación de ácido esteárico hasta oleico, lo que podría relacionarse con la farmación de sincitio y muerte celular ("'. La formación de sincitio parece estar muy relacionada con la depleción de los niveles de células CD4'("), y es posible que en el proceso de fusión celular, los cambios metabólicos registrados en las células huésped producirán la desaturación de los residuos de ácidos grasos de su membrana fosfalipídica. Así, la fluidez de las membranas puede ser necesaria para la integración y formación de sincitio(""'). La apolipoproteína Al (la principal de la fracción HDL) y sus análogos peptidico- hélice anfipáticos inhiben la forniación de sincitio VIHinducida"`,sugiriendo que la disminución de HDL, observada en la infección por VIH- 1, puede también contribuir a la generación de sincitio, favoreciendo, por lo tanto, la expresión de fenotipos del VIH-1. Así, las propiedades biológicas del VIH podrían estar influenciadas por las alteraciones del metabolismo lipídico`. 2.2 Niveles plasmáticos de lipoproteínas Las lipoproteínas plasmáticas pueden influenciar la función celular a través de cambios en la composición lipídica de la membrana celular resultando en transtorno de la inmunocompetencia del sistema humoral y celula?). Feingold et af') han sugerido que los disturbios del metabolismo de los triglicéridos observados en el paciente con SIDA, conducen a la aparición del fenotipo LDL - subclase B. Esta subclase de LDL está relacionada a otras variaciones en los niveles de lípidos y lipoproteínas, incluyendo elevaciones de triglicéridos, VLDL, y de lipoproteínas de densidad intermedia (lDL), así como bajas concentraciones de colesterol HDL, niveles altos de apolipoproteína B y niveles bajos de apolipoproteínaAl(". Grunfeld et aP' determinaron el grado de distribución del colesterol y de apolipoproteínas en las lipoproteínas plasmáticas de sujetos infectados con el VIH-1 y pacientes con SIDA. En ambos grupos de estudio encontraron niveles bajos de HDL y apoliproteína Al. En el grupo de pacientes con SIDA observaron niveles disminuidos de colesterol LDL, mientras que en el grupo de individuos infectados con VIH-1, una tendencia a la disminución en los niveles de colesterol LDL. Los niveles de apolipoproteína B-100 estuvieron disminuidos en ambos grupos lo que reflejaría aún más la disminución de los niveles de LDL("). Grunfeld y Feingold "') han propuesto que los niveles altos de VLDI, resultan de la lipólisis periférica, un disturbio metabólico observado en el Síndrome de Consumo asociado al SIDA. Dicha alteración moviliza loa ácidos grasos desde los depósitos tisulares lipídicos, los cuales son reesterificados hasta triglicéridos en el hígado y (1,12) luego vueltos a secretar al torrente sanguíneo . En este proceso, denominado "ciclo futil", los ácidos grasos son removidos del tejido adiposo y transportados hacia el hígado, y luego enviados de regreso al mismo tejido adiposo sin haber sido 12,13) utilizados como fuente de energía. (12,13).
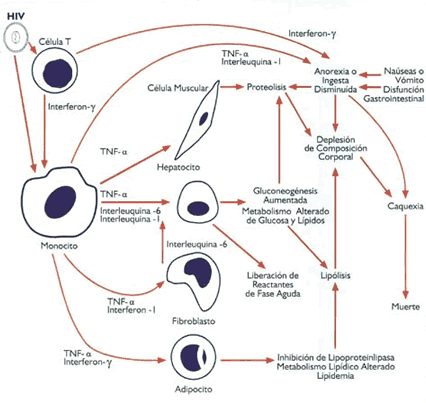
3. Mediadores de las alteraciones del Metabolismo de los Lípidos en la Infección por VIH-1 y SIDA 3.1 Citoquinas y Metabolismo Lipídico Las citoquinas son proteínas secretadas por los macrófagos y otras células del sistema inmunológica. Actúan como mensajeros biológicos y juegan un papel importante en la respuesta del sistema inmunológico contra las infecciones ("~ Los ejemplos más característicos lo constituyen la Interleuquina 6 (IL-6), Interleuquina l(IL-1), el Factor de Necrosis Tumoral alfa (TNF-ce) y los Interferones QFNs)("'"). De ellos, el TNF-u, IL-1, 117N -a e IFN - gamína han sido propuestos corno mediadores de los efectos de las infecciones sobre el metabolismo de los lípidos`. Tanto el TNF-a, como la IL-1 e IL-6 poseen actividad lipogénica, estimulando así la síntesis hepática de ácidos grasos (AG) al incrementar los niveles hepáticos de citrato, el cual constituye el principal activador alostérico de la Acetil CoA carboxilasa, la enzima limitante de la síntesis de AG ("'. Sin embargo, el IF`N-a posee una actividad estimulante directa de la síntesis hepática de AG, mecanismo que no tiene efecto sobre los niveles hepáticos de citrato"`. Las alteraciones causadas por las citoquinas han sido ampliamente descritas durante el curso la infección por VIH- 1 ( tanto a nivel de los órganos linfoides como en sangre periférica(", donde se han reportado niveles elevados de IFN-a, TNF-a, IL-6 e lL-l(`. Aunque las citoquinas parecen influenciar la replicación in vitro del VIH- 1 y están vinculadas a algunos eventos clínicos de la infección por VIH- l"'), todavía es difícil determinar una relación clara entre los niveles de citoquinas y los diferentes estadíos de la enfermedad (`. Zangerle et al han propuesto que las citoquinas tendrían relación con la disminución de los niveles de colesterol HDL durante la infección por VIH-l('). Sin embargo, estudios previos no demuestran diferencias significativas en los niveles de TNF en pacientes con SIDA(`), probablemente debido a su rápida (33,39) depuración de la circulación Las citoquinas también afectan la reesterificación de los ácidos grasos, un proceso en el cual las moléculas de ácidos grasos y el glicerol reaccionan reversiblemente para producir triglicéridos(12,41) . Los ácidos grasos son movilizados desde la periferia y luego procesados nuevamente en el hígado hasta triglicéridos. Este proceso, además de la lipogénesis hepática de novo, originaría un incremento de los niveles de VLDL, lo que en la infección por VIH-1- contribuye a la hipertrigliceridemia observada". Las concentraciones bajas de TNF-a causan aumento de los niveles de VLDL, mientras que las concentraciones altas causan tanto producción elevada de VLDL, como disminución de la actividad de la lipoproteini ¡pasa `," perjudicando la depuración y captación de los triglicéridos disponibles y generándose, como consecuencia, una persistente hipertrigliceridemia (12,40) . La incorporacion de triglicéridos y su empaque dentro de los adipocitos encuentra también inhibida, contribuyendo así a los niveles ascendentes de triglicéridos circulantes ("). Los interferones - <x , -13 y -gamma pueden inducir resistencia de la célula huésped frente a una gran variedad de infecciones virales incluyendo los retrovirus. Estudios in vitro han reportado una actividad antiviral sinergista entre el IFN-gamma y el TNF- oi ("). El IFN -a estimula la lipogénesis hepática in vivo y disminuye el almacenaje de lípidos en las células adiposas cultivadas in vitro. Ambos mecanismos están relacionados con la (3 hipertrigliceridemia observada en las infecciones . Estudios adicionales sugieren que el lFN-oi, humano podría ser más efectivo en reducir la replicación in vitro de VIH-1 e IFN-gamma"), y que el IFN-a puede modular el metabolismo lipídico in vivo e in vitro, lo que contribuirá a la hipertrigliceridemia observada en el SIDA (39) . En algunas circunstancias, tales como el ayuno, existe una correlación entre los niveles de IFN-(x, la disminución de la depuración de triglicéridos y el incremento de la síntesis hepática de ácidos grasos (12). Grunfeld et al han reportado que la hipertrigliceridemia en el SIDA se relaciona con los niveles circulantes de IFN-a, y no con los de TNF-(xni tampoco con los de IL-1. Dado que los efectos sobre el metabolismo de los lípidos son sostenidos, aún después de ser depurados del plasma (32,39), sus resultados sugieren que los niveles circulantes sistémicos de TNF-(x e IL-1 pueden no ser requeridos para producir cambios metabólicos sistémicos (39). Niveles bajos de colesterol, sin embargo, parecen preceder al aumento de los niveles de triglicéridos y no se correlacionan con los de IFN (12) 3.2 Citoquinas, Metabolismo Lipídico y Síndrome de Consumo La hiperactivación del sistema inmunológico y el disbalance de las citoquinas juegan un rol importante en el Síndrome de Consumo, entidad frecuentemente observada en el curso del SIDA(12, 13). Los posibles mediadores del consumo incluyen la baja ingesta calórica debido a la anorexia, la incapacidad para absorber los nutrientes ingeridos`, y los estados de (12,44) hipermetabolismo y estrés oxidativo . La causa del consumo en la infección por VIII- 1 es multifactorial y se ilustra en la Figura l(45). Durante el estrés catabólico, inducido por el VIII- 1, las citoquinas (e.g. IF-gamina, TNF-a, IL-1) son liberadas de la interacción del virus con las células del sistema inmunológico o en presencia de cualquier infección sevcundaria (e.g. de rigen gastrointestinal) (13,45). In vitro, el TNF-a disminuye la actividad de la lipoproteinlipasa e induce un estado de hipertrigliceridemia al promover el catabolismo del tejido adiposo, un efecto que también es compartido por otras citoquinas (12,39,47) . Además, las citoquinas pueden incrementar los niveles plasmáticos de triglicéridos al estimular la lipogénesis hepática y la producción de VLDL, así como también la síntesis hepática y movilización periférica de los ácidos grasos libreS(3,12) . Los AGL movilizados serán reesterificados hasta triglicéridos en el hígado y luego vueltos a secretar, contribuyendo al incremento de los niveles plasmáticos de VLDL. El resultado es un mecanismo al que se le ha dado el nombre de "ciclo futiP, en el cual los AGL son transportados desde el tejido adiposo hasta el hígado y devueltos nuevamente al tejido adiposo sin haber sido utilizados como fuente de energíaT"'. La síntesis inapropiada de lípidos, ftente a la pérdida de peso corporal, podría conducir a la pérdida desproporcionada de masa magra relativa a los depósitos lipídicos, contribuyendo así al estado de hipermetabolismo (1). Las citoquinas interfieren con los procesos metabólicos en el hígado lo que genera trastornos del metabolismo glucídico (e.g_ incremento de la gluconeogenesis), de las proteínas (proteolisis) y de los lípidos (lipólisis y pérdida de las reservas corporales de grasay1 12.45,46'. Los resultados finales de estos mecanismos combinados son, nuevamente, anorexia, ingesta inadecuada de nutrientes, depleción de la composición corporal, consumo y muerte del organismo huésped (45). Resumen Los niveles plasmáticos anormales de lípidos y lipoproteínas han sido reportados en estudios realizados en individuos sintomáticos y asintomáticos infectados con el VIH-1. La hipocolesterolemia e hipertrigliceridemia son los disturbios más frecuentemente observados en las diferentes fases de la progresión de la enfermedad. Estos cambios en el perfil lípidico se encuentran asociados a la activación viral y la consecuente respuesta inimine, en especial, la producción de citoquinas. El estado del metabolismo de los lípidos puede ser aún más influenciado por enfermedades oportunistas y la anorexia. En consecuencia, la alteración del metabolismo lipídico podría contribuir a la progresión de la enfermedad, el síndrome de consumo y la elevada mortalidad frecuentemente observada en la infección por VIII- 1.
Bibliografía 1. Hallerstein M, Grunfeld C, Wu K. Increased De Novo Hepatic lipogenesis in Human Immunodeficiency Virus Infection. J Clin Endocrinol and Metab 1993; 76(3): 559-565. 2. Grunfeld C, Kotler D, Hamadeh R. Hypertriglyceridernia in the Acquired Immunodeficiency Syndrome. Am J ofmed 1989; 86: 27-31 3. Gallin J, Kaye D, O'Leary W. Serum Lipds in Infection. NEng1JMed 1969; 281:1081-86 4. Edginton T, Curtiss L. Plasma Lipoproteins with Bioregulatory Properties including the Capacity to Regulate Lymphocyte Function and the Immune Response. CancerResl981,41:3786-3788. 5. Flegel W, Woepl A, Manuel D. Inhibition of Endotoxin-Induced Activation offluman Monocytes by Human Lipoproteins. Infection and immunity 1989; 57: 2237-2245. 6. Peck M, Mantero-Atienza E, Miguez-Burbano M. The Esterified Plasma Fatty Acid Profile is Altered in EarlyHIV-1 Infection. Lipids 1993; 28:593-597. 7. Shor-Posner G, Basit A, Lu Y. Hypocholesterolemia is Associated with Immine Dysfilnetion in Early HIV-1 infection. Am Jofmed 1993; 94: 515-519.. 8. Zangerle 11, Sarclettl M, Gallati H. Decreased Plasma Concentrations of HDI, Cholesterol in HIV-Infected Individuals are Associated with Immune Activation. J of Acquired Immune Deficiency Syndromes 1994; 7:1149-1156. 9. Christeff N, Lortholary 0, Casassus P. Serum Lipid Concentrations with Reference to the Clinical and Immunological Status of HIV Infected Men. Annales de Medecine Interne 1995. 146(7): 490 - 495. 10. Lees R, Fiser R, Beisel W. Effects of an Experimental Viral Infection on Plasma Lipid and Lipoprotein Metabolism. Metabolism 1972; 21: 825-833. 11. Sammalkorpi K, Valtonen V, Kerttula Y. Changes in serum lipoprotein pattern induced by acute infections. Metabolism 1988; 37:859-865. 12. Grunfeld C, Feingold K. Metabolic Disturbances and Wasting in the Acquired Immunodeficiency Syndrome. NEnglJMed 1992; 327 (5):329 - 335. 13. Grunfeld C, Feingold K. The role of the Cytokines, Interferon Alpha and Tumor Necrosis Factor in the Hypertriglyceridemia and Wasting of AIDS. J Nutr 1992; 122:749-753. 14. Baum MK, Shor-Posner G, Lu Y. Normal Triglyceride Levels in Early HIV-1 Infection. AIDS 1994,8(l):131132. 15. Rovirs E, Belda A, Martinez M. The Prognostic Value of Plasma Triglycerides in Human Immunodeficiency Virus Infection. Revista Clinica Espahola 1995; 195(7): 455-458. 16. Douvas G, May M, Pearson R. Hypertriglyceridemic Serum, Very Low Density Lipoprotein and Iron Enhance Mycobacterium Avium Replication in Human Macrophages. J of infectious Diseases 1994- 170:12481255 17.Gonzales-Clemente J, Miro J, Navarro M. High Triglyceride Levels as a Predictor of Mortality in AIDS Patients. AIDS 1993; 7(7): 1022-1023. 18. Shor-Posner G, Zhang G, Campa A, GutierrezRodriguez R. Hypertriglyceridemia Mortality and Cardiac Function in Pediatric AIDS (inpress). 19.Grunfeld C, Pang M, DoerrIer W. Lipids, Lipoprotems, Triglyceride Clearance and Cytokines in Human Inummodeficiency Virus Infection and the Acquired Immunodeficiency Syndrome. J Clin EndocrinolandMetab 1992; 74(5):1045-1052. 20. Klein A, Mercure L, Gordon P. The Effect of HIV Infection on the Lipid Fatty Acid Content in the Membrane of Cultured Lymphocytes. AIDS 1990, 4:865-867. 21.Lynn W, Tweedale A, Cloyd M. Human Immunodeficiency Virus (HIV-1) Cytotoxicity: Perturbation of the Cell Membrane and Depression of Phospholipid Synthesis. Virology 1988; 163:43-51. 22. Klein A, Bruser M, Bast M. Progress ofHIV Infection and Changes in the Lipid Membrane Structure of CD4+ cells. AIDS 1992; 6(3): 332-333 23. Boucher A, Lange J, Miedema F. HIV-1 Biological Phenotype and the Development of Zidovudine Resistance in Relation to Disease Progression In Asymptornatic Individuals During Treatment. AIDS 1992; 6:1259-1264 24. Riibsamen-Waigmann H, Schr6der B, Biesert L. Markers for HIV-disease Progression in Untreated Patients and Patients Receiving AZT: Evaluation of Viral Activity, AZT Resistance, Serum Cholesterol, 2microglobulin, CD4+ cell counts and HIV antigen. Infection 1991- 19:S77-S82. 25. Minggao S, Taylor J, Fahey J. Early Levels of C134, Neopterin, and 2-microglobulin Indicate Future Disease Progression. JofClin Immunol 1997; 17(l):43-52. 26. Melmed P, Taylor J, Detels RL Serum Neopterin Changes in HIV-Infected Subjects: Indicator of Significant Pathology, CD4'T cell Changes, and the Development of AIDS. JofAcquiredImmDeficSyndr 1989;2(l):70-76 26. Phillips A, Sabin C, Elford J. Serum 2-M'Croglobulin at HIV-1 Seroconversion as a Predictor of Severe Immunodeficiency Dunng Ten Years of Follow - up. J of Acquired Imm DeJic Syndr and Human Retrovirol 1996; 13 (3):262 -266. 28. Begin M, Manku M, Horrobin D. Plasma Fatty Acid Levels in Patients with AIDS and in controls. Prostagl LeukotrandEssentFattyAcids 1989.37:135-137. 29. Apostolov K, Barker W, Galpin S. Syncytia formation in HIV infected cells is associated with an increase in cellular oleic acid. Federation of European Biochemical Societies 1989-,250 (2): 241-244. 30. Ownes R, Anantharamaiah G, Kahlon J. Apolipoprotein Al and its amphipatic helix peptide analogues inhibit HIV syncytium formation. J Clin Invest 1990; 86: 1142-1150. 31. Feingold K, Krauss R, Pang M. The Hypertriglyceridemia of Acquired Immunodeficiency Syndrome is Associated with an Increased Prevalence of Low Density Lipoprotein Subclass Pattern B. J Clin EndocrinolandMetab 1993.76(6):1423-1427 32. Shizou A, Hirano T, Taga T. Biology of multifunctional cytokines: IL-6 and related molecules (IL-I and TNF).FASEBJournaL 1990.4:2860-2867. 33. Grunfeld C, Adi S, Soued M. Search for mediators of the lipogenic effects of Tumor Necrosis Factor: Potential role of Interleukin 6. Cancer Research 1990. 50: 42334238. 34. Grunfeld C, Soued M, Adi S. Evidence for two classes of cytokine that stimulate hepatic lipogenesis: Relationships among tumor necrosis factor, interleukinI and interferon-alpha. Endocrinology 1990. 127: 4654. 35. Fauci AS. Multifactorial nature of human immunodeficiency virus disease: implications for therapy. Science 1993,262: 1011-1018. 36. Yamamoto N. The Role of cytokines in the acquired immunodeficiency syndrome. IntJClinLahRes.1995, 25:29-34. 37. Matsuyama T, Kobayashi N, Yamamoto N. Cytokines and HIV infection: Is AIDS a tumor necrosis factor disease? AIDS 1991,5:1405-1417. 38. Rimaniol AC, Zylberberg H, Zavala F. Inflammatory cytokines and inhibitors in HIV infection: correlation between interleukin- I receptor agonist and weight loss. AIDS 1996,10:1349-1356. 39. Grunfeld C, Kotler D, Shigenaga JK. Circulating Interferon-ot levels and hypertriglyceridernia in the acquired immunodeficiency syndrome. The Am J of Med 1991. 90:154-16 1. 40. Feingold K, Serio M, Adi S. Tumor Necrosis Factor stimulated hepatic lipid synthesis and secretion. Endocrinology. 1989; 124:2336-2342. 41. BeHSJ,Mascio]iEA,ForseRA. Nutrition support and the human immunodeficieney virus (HIV). Parasitology. 1993; 107: S53-S67. 42. Wong GH, Goeddel DV. Tumor Necrosis Factor cc and inhibit virus replication and synergize with interferons. Nature 1986.323: 819-822. 43. Hartshorn KL, Neumeyer D, Vogt MW. Activity of interferon alpha, beta and gamma against HIV replication in vitro,AIDSRes Human Retroviruses 1987. 3:125-133. 44. Baldwin C. Nutrition and HIV Infection. 2nd Ann Conf on HIV and Nutrition. London, Oct. 1993. 45. TchekmedyianNS. Clinical Approaches to Nutritional Support in Cancer. Current Opinion in Oncology 1993; 5:633-638. 46. Moldawer L, Rogy M, Lowry S. The Role of Cytokines in Cancer Cachexia. JParent Enter Nutr 1992; 16:43S49S. 47. Grinspoon S, Bilezikian J. HIV Disease and the Endocrine System. N Engl J Med 1992. 327: 13601365.
(*) Fogarty International AIDS Research
Training Program, University ofMiami School ofMedicine. Centerfor Disease
Prevention. Profesor Auxiliar de Bioquimica y Nutrici6n. Facultad de Medicina
"Hip6lito Unanue ", UniversidadNacional "Federico Villarreal". |