|
|
||||
|
La hélice dorada y la medicina molecular Resumen La práctica médica se beneficiará enormemente de los desarrollos recientes en biología molecular y en la tecnología del ADN recombinante. En la presente revisión se presentan algunas de las contribuciones más importantes en biología molecular en el contexto de su aplicación potencial en la investigación bio-médica y la práctica clínica . Summary The medical practice will benefit enormously from current developments in molecular biology and DNA recombinant technology. In the present review some scientific and technological milestones in molecular biology are presented in relation to the potential application in the bio-medical research and clinical practice. Cuando en 1953, Francis Crick, un maduro estudiante inglés de doctorado, a la sazón con 37 años de edad y James Watson, un joven talentoso post-doc americano de 25 años, publicaron en la revista científica británica Nature la estructura helicoidal del ADN (1) no imaginaron que 50 años en adelante la secuencia completa del genoma humano, compuesto de tres mil millones de pares de bases estaría almacenada en super-computadoras y a disposición de los ciudadanos de la tierra vía Internet. Lo que probablemente no ocupaba las aspiraciones de nadie entonces, constituye actualmente un monumental esfuerzo internacional con el objetivo de descifrar toda la información genética almacenada en la doble hélice de ADN del núcleo de las células humanas. Señalar la secuencia de eventos desde aquel verano del 53 hasta nuestros días es una tarea demasiado extensa para los confines del presente artículo. Sin embargo, apuntar algunos de los descubrimientos importantes en Biología Molecular es un ejercicio útil que permite resaltar la fecunda relación entre la Biología Molecular y la Medicina. La Hélice doble del ADN
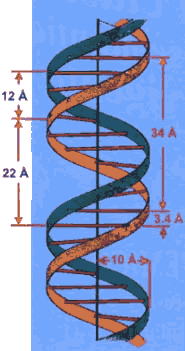
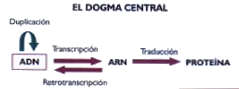
Los patrones de difracción de rayos X obtenidos de cristales de ADN y la construcción de modelos moleculares a escala permitieron dilucidar la estructura del ADN (1). El ADN es un polímero ácido constituido por unidades químicas denominados nucleotidos que se unen entre sí por enlaces fosfodiester. Los nucleotidos están conformados por una base nitrogenada que puede ser Adenina, Guanina, Citosina o Timina, una azúcar de cinco carbonos desoxiribosa a la que se une un grupo fosfato. El ADN tiene una estructura de doble hélice formada por dos cadenas independientes y complementarias. La complementariedad de las cadenas de ADN en la doble hélice es el producto de un orden de interacciones estables entre los nucleotidos: Guanina interactúa con Citosina por tres enlaces de hidrógeno y Adenina con Timina por dos enlaces de hidrógeno. Cualquier violación de este orden de interacciones es detectada por el sistema celular de reparación del ADN que reemplaza el nucleotido discordante por el complementario. La estabilidad de la estructura de doble hélice del ADN es notable y conlleva a la existencia de moléculas grandes, probablemente las mayores que se hayan dado en la naturaleza, por ejemplo el cromosoma 1 humano es una molécula de ADN lineal que tiene una longitud de 250 millones de pares de bases (bp). Una propiedad fundamental de la doble hélice es la complementariedad de las cadenas del ADN, la cual permite el almacenamiento de la información genética, su transmisión entre generaciones por replicación y su mantenimiento ya que una hebra siempre sirve de plantilla para reparar la complementaria dañada. Figura 1 . Dogma Central y Código Genético Lo que ha venido a conocerse como el Dogma Central de la Biología Molecular establece que la información genética se almacena y perpetúa como secuencias de ácidos nucleicos y la información se ejecuta en funciones realizadas por proteínas. Replicación es el proceso que perpetúa la información genética entre generaciones, y Expresión Genética se refiere a los eventos celulares que conducen a la ejecución de la información almacenada en el ADN en dos procesos centrales: 1. Transcripción es el proceso de síntesis de los ARN mensajeros (mRNAs) que portan la información de los genes codificados en el ADN con instrucciones para la síntesis de las correspondientes proteínas y 2. Traducción es el proceso de síntesis de proteínas en los ribosomas sobre la plantilla de instrucciones que porta mRNA. Figura 2. La unidad de información genética es el codon- una secuencia de tres nucleotidos que codifica la incorporación de un amino ácido durante el proceso de traducción, además hay codones que señalan el inicio y el termino del mensaje. La cartilla de codones y su respectivos amino ácidos es lo que se denomina el Código Genético. La universalidad del código genético se refiere, a que con muy pocas excepciones, es el códice que permite a los ribosomas traducir la información genética en proteínas de toda la gama de organismos aún tan distantes evolutivamente como archeabacterias y humanos.
Tecnología del ADN recombinante Las herramientas moleculares necesarias para
la manipulación de los genes alcanzaron un alto grado de difusión gracias
a su disponibilidad comercial. Actualmente, diversas empresas biotecnológicas
ofrecen decenas de diferentes enzimas de restricción, que son endonucleasas
que escinden el ADN en secuencias específicas, ADN ligasas que son enzimas
que unen moléculas de ADN restituyendo los enlaces fosfodiester mediante
la hidrólisis de ATP, y ADN polimerasas que duplican el ADN a partir de
los precursores nucleotidos trifosfato. El uso combinado de estas enzimas
constituye el fundamento de la tecnología del ADN recombinante, i.e. escindir
un gen de su entorno natural en el cromosoma, unir el gen escindido a
un fragmento de ADN diferente y replicarlo en varios órdenes de magnitud
mediante la acción de la ADN polimerasa o por clonación en un sistema
vector/hospedador. Productos conspicuos de la tecnología del ADN recombinante
son las proteínas recombinantes generadas en microorganismos, células
en cultivo o en animales transgénicos. Las denominadas proteínas recombinantes
humanas de primera generación son producidas en bacteria, levaduras o
en células humanas en cultivo. Algunos ejemplos de productos de primera
generación se muestran en la tabla 1.
Las proteínas humanas recombinantes de segunda generación se caracterizan por ser proteínas recombinantes modificadas por fusión, mutación o deleción de determinadas regiones de su estructura proteíca para potenciar su acción farmacológica. Se vislumbra que la tercera generación de proteínas recombinantes serán producidas en tejidos humanos después de la transferencia de los genes relevantes (2) al genoma del paciente. La segunda y tercera generación de estos productos farmacéuticos se encuentran en desarrollo. Una de las más brillantes innovaciones en la tecnología del ADN recombinante es la amplificación del ADN por la reacción en cadena de la polimerasa (PCR). La reacción de amplificación en cadena consiste en ciclos repetidos de replicación de una secuencia de ADN determinada por la acción de ADN polimerasas resistentes a altas temperaturas (95°C). PCR puede amplificar una sola molécula de ADN millones de veces (3). Debido a su alta sensibilidad y especificidad, PCR es ampliamente aplicado en patología molecular principalmente en la detección de células transformadas y agentes patógenos (4). Bibliotecas genéticas Una biblioteca genética constituye la colección de fragmentos de las secuencias del ADN del genoma de un organismo. Su construcción requiere de la fragmentación de las cadenas largas de ADN por la acción de una enzima de restricción, estos fragmentos son mantenidos de modo recombinante en E. coli o levadura. Cada célula de E. coli o levadura mantiene un único fragmento de ADN clonado en un vector, de tal modo que en una población de estas células en cultivo (en el rango de 107 a 108 células) se puede contar con todos las secuencias del genoma del organismo donde cada fragmento es portado por cada microorganismo recombinante. La población de células que contiene todos los fragmentos de ADN recombinante de un genoma se denomina la biblioteca genética del organismo. Una variante de las bibliotecas genéticas son las llamadas bibliotecas de expresión las cuales contienen solamente los genes que están siendo expresados en un tejido humano particular. Estas bibliotecas son construidas sobre la base de la población de mRNAs sintetizados en el tejido. La construcción de la biblioteca de expresión requiere del uso de la transcriptasa reversa que retrotranscribe los mRNA a moléculas de cADN- denotado así para indicar su procedencia. Existen en la actualidad bibliotecas de expresión provenientes de tejidos humanos en diferentes estadíos de desarrollo y en condiciones patológicas como el cáncer. Las bibliotecas de expresión son la materia prima para la clonación de los genes humanos y su producción como proteínas humanas recombinantes. Vectores y Hospedadores La construcción de la bibliotecas genéticas requiere de vectores y hospedadores. Los vectores son unidades de ADN que se autoreplican en un microorganismo o células humanas y los cuales pueden portar fragmentos de ADN de origen diverso. El vector debe contar con los elementos que le permitan ser replicados y mantenidos por la maquinaria celular hospedadora. Los vectores son propagados en hospedadores que pueden ser células de E. coli, levadura (S. cerevisiae) o células mamíferas. Los vectores pueden ser estructuras sencillas como plásmidos o muy complejos como los cromosomas artificiales de levadura (YAC) (5) y los cromosomas artificiales mamíferos (MAC) (6). Como puede apreciarse en la tabla 2, los vectores tienen diferente capacidad de portar longitudes variables de secuencias de ADN. Los cromosomas artificiales mamíferos se vislumbran como vectores de terapia genética de condiciones causada por genes mayores como el de la distrofina, cuya disfunción causa la distrofia muscular de Duchenne-Becker, gen que tiene una longitud de secuencia de 2.4 Mb (7).
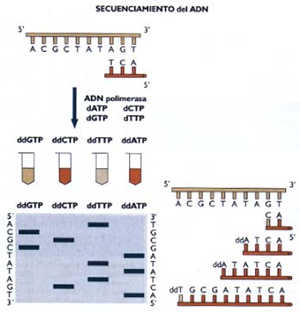
Secuenciamiento del ADN El método de Sanger permite obtener la secuencia
de nucleotidos de un fragmento de ADN de tamaño de 600-1000 bp (8). El
secuenciamiento requiere de la acción de la ADN polimerasa que sintetiza
la cadena de ADN en presencia de nucleotidos trifosfato (dNTPs) y de análogos
de los nucleotidos trifosfato (ddNTPs) acoplados a un compuesto radioactivo
o cromogénico. Los ddNTPs interrumpen el proceso de síntesis del ADN ya
que no permiten la elongación de la cadena sintetizada. La reacción de
polimerización en estas condiciones genera fragmentos de ADN de tamaño
variable, los cuales son separados por electroforesis y visualizados.
Como se muestra en la figura 3. El esfuerzo por descifrar
la secuencia de ADN de las 3 x 109 bp del genoma humano impulsó la automatización
y robotización del procedimiento de Sanger (9). Para tener una idea de
la magnitud de la tarea de secuenciar el genoma humano, tomemos como ejemplo
el esfuerzo requerido para obtener la secuencia de ADN del cromosoma 22
recientemente publicada (10). La secuencia obtenida es de 33.4 millones
de pares de bases de la región cromosomal que contiene la mayoría de los
genes del cromosoma 22. Considerando la capacidad actual de secuenciar
ADN, un solo secuenciador automático generando 10,000 bp de secuencia
cada 24 horas y trabajando ininterrumpidamente completaría la secuencia
del cromosoma 22 en 18 años, y en secuenciar todo el genoma humano, cien
veces el tamaño del cromosoma 22, tardaría 18 años x 100 = 1800 años.
La secuencia del cromosoma 22 ha sido obtenida en aproximadamente cinco
años gracias a la participación de un consorcio de grupos de investigación.
Testimonio del esfuerzo conjunto son los doscientos y tantos investigadores
que tienen autoría en la publicación (10).
El secuenciamiento del ADN se usa rutinariamente en la determinación de la identidad humana ya que permite tanto la identificación de las personas, como la determinación de la filiación familiar Análisis Seriado de la Expresión Genética (SAGE) y chips de ADN La identificación de los genes que tienen una patrón de expresión alterado en condiciones patológicas es importante para el entendimiento de la bases moleculares de la patogénesis de una condición particular y para el diseño de una potencial intervención terapéutica. Dos tecnologías desarrolladas recientemente permiten determinar variaciones en los patrones de expresión de virtualmente todos los genes de un tejido. La primera es el Análisis Seriado de la Expresión Genética (SAGE) que detecta secuencias cortas de ADN (10 bp) que contienen información suficiente y sirven como un código de barras para identificar y cuantificar la expresión de genes en un tejido particular (11). La segunda, son los chips de ADN que consisten en pequeñas láminas de plástico o silicio sobre cuya superficie se han inmovilizado o sintetizado miles de diferentes fragmentos de And (12, 13). La totalidad de genes humanos, es decir de 60-80,000 genes pueden inmovilizarse en un sólo chip de 1 cm2. La expresión genética es determinada en los chips mediante hibridación de ADN: La sonda de ADN que contiene un fluorocromo detecta secuencias específicas entre las miles de secuencias inmovilizadas en el chip y la intensidad de la hibridación es detectada por un láser, visualizada por microscopía confocal y la imagen procesada en una computadora. La intensidad de la señal es proporcional a la abundancia de la expresión de un gen particular. Los chips de ADN permiten el análisis de decenas de miles de secuencias en un sólo ensayo. Aunque las aplicaciones diagnósticas de los chips de ADN se encuentran a nivel de prototipo se vislumbra su utilidad en a) la detección casi inmediata de todas las mutaciones en el genoma de un individuo, sean estas alteraciones en la secuencia, o el número de copias de genes, b) la detección de miles de agentes patógenos en un solo ensayo y c) prognóstico de un individuo a desarrollar enfermedades genéticas multifactoriales, es decir causadas por la disfunción de más de un gen, por ejemplo asma, diabetes, entre otras. Animales Transgénicos Los animales transgénicos son portadores de genes no propios de su especie, los cuales han sido incorporados en su genoma mediante tecnología de ADN recombinante. La incorporación de estos genes foráneos en el genoma recipiente es posible gracias a la propiedad que tienen las moléculas de ADN por recombinarse. Recombinación es el procedimiento normal de intercambio de segmentos de secuencia entre moléculas de ADN. Los primeros animales transgénicos en obtenerse fueron ratones. Estos se lograron mediante la inyección de un gen foráneo al núcleo de células embrionarias totipotentes. El ADN foráneo se incorpora inmediatamente en el genoma de la célula embrionaria de modo específico por recombinación con una frecuencia muy baja (˜ 10-7). Las células así transformadas son incorporadas en el blastocisto en desarrollo y luego implantados en el útero de ratonas recipientes donde completan su desarrollo. La progenie de ratones de la primaria generación son ratones quiméricos debido a que portan células con diferente contenido genético i.e. tejidos que provienen de células del blastocisto receptor y tejidos transgénicos que provienen de las células que han incorporado el gen foráneo en su genoma. Si las células transformadas forman parte del tejido germinal, la subsecuente cruza de ratones quiméricos con ratones normales produce una línea de ratones transgénicos heterocigotos y eventualmente la cruza de transgénicos heterocigotos llega a producir una línea transgénica homocigota (14). Los animales transgénicos han permitido contar con una gama más amplia de modelos animales de enfermedades humanas, que en el pasado sólo se podían obtener por las mutaciones espontaneas que ocurrían en las líneas de ratones de laboratorio. Animales clonados: Tecnología de la transferencia nuclear La obtención de animales clonados a partir de células diferenciadas adultas (15) ha sido posible gracias a la tecnología de transferencia nuclear. El procedimiento consiste en la transferencia del núcleo que contiene el material genético de una célula donadora al citoplasma de un oocito o cigoto al cual se le ha quitado núcleo. El oocito sin núcleo contribuye con los factores citoplasmáticos necesarios para iniciar el completar el desarrollo embrionario hasta un individuo adulto clonado. La gama de animales clonados por transferencia nuclear incluyen ovejas (15) ratones (16), cabras (17) y vacas (18). La aplicación conjunta de transgenia y transferencia nuclear (19) constituye una gran promesa para medicina del futuro ya que permitirá la producción de una variedad de hormonas, factores y otras proteínas humanas en animales, así como la generación de órganos humanos en animales transgénicos en los cuales los marcadores de histocompatibilidad han sido modificados por ingeniería genética. La Hélice Dorada y la Medicina Molecular La obtención de la secuencia total del ADN nuclear es el objetivo central del proyecto del genoma humano. El genoma haploide humano tiene un contenido de ADN de 3 x 109 bp distribuido en 22 cromosomas autosómicos y 1 cromosoma sexual. El proyecto del genoma humano tiene un enorme potencial en la investigación y la práctica médica. La información de la secuencia de todos los genes humanos unida a la tecnología del ADN recombinante tiene aplicaciones en virtualmente todas las ramas de la práctica médica, sea por la producción de un amplio rango de agentes terapéuticos y de diagnóstico, por permitirnos entender aspectos de la biología humana y por descubrir las causas de enfermedades como la hipertensión, enfermedad cardiaca, desórdenes psiquiátricos y el cáncer, entre otras. Al caminar por Portugal Place en Cambridge, no escapaba a mi atención la hélice dorada que Francis Crick hizo colocar en la fachada de su casa de entonces, como una celebración personal al desciframiento de la estructura de doble hélice del ADN. En cierto modo la finalización del proyecto del genoma humano en el año 2003 -coincidente con el 50avo aniversario del descubrimiento de la estructura del And (20)- cierra un ciclo y nos endosa un legado que nos fortalece en el esfuerzo por disminuir la miseria causada por las enfermedades que aquejan a la humanidad. Bibliografía 1. Watson, JD. and Crick, FHC. A structure of desoxyribosenucleic acid. Nature 1953; 171: 737 2. Buckel, P. Recombinant proteins for therapy. Trends in Pharmacological Science 1996; 17: 450-456. 3. Saiki, RK; Gelfand, DH; Stoffel, S; Scharf, SJ; Higuchi, R; Horn, GT; Mullis, KB. and Erlich, HA. Primer-directed enzymatic amplification of DNA with a thermostable DNA polymerase. Science 1988; 239: 487- 491. 4. Going, JJ. and Gusterson, BA. Molecular pathology and future developments. European Journal of Cancer 1999; 35:1895-1904. 5. Burke, DT; Carle, GF. and Olson, MV. Cloning of large segments of exogenous DNA into yeast by means of artificial chromosome vectors. Science 1987; 236: 806- 812. 6. Larin, Z. and Monaco, AP. YACs. BACs, PACs and MACs: artificial chromosomes as a research tools. Trends in Biotechnology 1994; 12: 280-286. 7. Huxley, C. Mammalian artificial chromosomes: a new tool for gene therapy. Gene Therapy 1994; 1: 7-12. 8. Sanger, F; Nicklen, S. and Coulson, AR. DNA sequencing with chain terminating inhibitors. Proceedings of the National Academy of Science 1977; 74: 5463-5467. 9. Hunkapiller, T; Kaiser, RJ; Koop, BF. and Hood, L. Large-scale and automated DNA sequence determination. Science 1991; 254: 59- 67. 10. Dunham, I; Shimizu, N; Roe, BA; Chissoe, S; Hunt, AR; Collins, JE; Bruskiewich, R; Beare, DM; Clamp, M; Smink, LJ; Ainscough, R; Almeida, JP. et al. The DNA sequence of human chromosome 22. Nature 1999; 402: 489-495. 11. Velculescu, VE; Zhan, L; Vogelstein, B. and Kinzler, K. Serial analysis of gene expression. Science 1995; 270: 484 - 487. 12. Fodor, SP; Rava, RP; Huang, XC; Pease, AC; Holmes, CP. and Adams CL. Multiplexed biochemical assays with biological chips. Nature 1993; 364: 555-556 13. Schena, M; Shalon, D; Davis, RW. and Brown, PO. Quantitative monitoring of gene expression patterns with a complementary DNA microarray. Science 1995; 270: 467- 470. 14. Capecchi, MR. Altering the genome by homologous recombination. Science 1989; 244: 1288-1292. 15. Wilmut, I; Schnieke, AE; McWhir, J; Kind, AJ. and Campbell, HS. Viable offspring derived from fetal and adult mammalian cells. Nature 1997; 385: 810-813. Scnieke, AE; Kind, AJ; Ritchie, WA; Mycock, K; Scott, AR; Ritchie, M; Wilmut, I; Colman, A. and Campbell, KHS. Human Factor IX Transgenic sheep produced by transfer of nuclei from transfected fetal fibroblasts. Science 1997; 278: 2130-2133. 16.Wakayama, T; Rodriguez, I; Perry, AC; Yanagimachi, R. and Mombaerts, P. Mice cloned from embryonic stem cells. Proceedings of the National Academy of Science USA 1999; 96:14984-14989. 17. Baguisi, A; Behboodi, E; Melican, DT; Pollock, JS; Destrempes, MM; Cammuso C, Williams, JL; Nims, SD; Porter, CA; Midura, P; Palacios, MJ; Ayres, SL; Denniston, RS; Hayes, ML; Ziomek, CA; Meade, HM; Godke, RA; Gavin, WG; Overstrom, EW; Echelard, Y. Production of goats by somatic cell nuclear transfer. Nature Biotechnology 1999; 17: 456 - 461. 18. Kubota, C; Yamakuchi, H; Todoroki, J; Mizoshita, K, Tabara; N, Barber, M. and Yang, X. Six cloned calves produced from adult fibroblast cells after long-term culture. Proceedings of the National Academy of Science USA 1999; 97: 990-995. 19. Acnieke, AE; Kind, AJ; Ritchie, WA; Mycock, K; Scott, AR; Ritchie, M; Wilmut, I; Colman, A. and Campbell, KHS. Human Factor IX Transgenic sheep produced by transfer of a nuclei from transfected fetal fibroblast. Science 1997; 278:2130-2133 20. Collins, FS; Patrinos, A; Jordan, E; Chakravarti, A; Gesteland, R. and Walters, L. (1998) New goals for the U.S. Human Genome Project: 1998-2003. Science 2000; 282: 682-689. (*) Profesor Principal
de Bioquímica y Biología Molecular, |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||