|
|
||||
|
El ADN y el diagnóstico molecular de enfermedades infecciosas Dr. Jorge Arévalo Zelada (*) Resumen La Biología Molecular ha contribuido no solamente al mejor entendimiento de las enfermedades infecciosas, sino que también ha proporcionado herramientas moleculares para el diagnóstico. Es posible mediante ella, detectar genomas de patógenos en muestras de biopsia o fluidos de pacientes. Debido a la elevada afinidad de las cadenas complementarias del ADN, es posible detectar muy pocos organismos en una muestra a ser analizada. Dos principales metodologías son presentadas en este reporte: La hibridación con sondas de ADN y la Reacción en Cadena de la Polimerasa. Esta última es el método de detección más sensible. Una lista resumen de los kits diagnósticos para diferentes enfermedades es presentada al final del artículo. Summary Molecular Biology has contributed not only to a better understanding of infectious diseases but it has also provided molecular tools for diagnosis. Using these tools it is possible to detect pathogen genomes in biopsy samples or in body fluids from patients. Because of the high affinity of complementary DNA strands (10-23 M), it is possible to detect very few organisms within a sample to be tested. Two main methodologies are presented in this report: Hybridization with DNA probes and the Polymerase Chain Reaction. The latter is the most sensitive detection method. A summary list of the available diagnostic kits for different diseases is presented at the end of this article. Un ciudadano europeo se percata de una úlcera pequeńa localizada en su mano derecha, poco tiempo después de haberse clavado una astilla cuando realizaba trabajos de carpintería doméstica. En un inicio no se preocupó, pero la herida fue incrementándose muy lentamente durante el último ańo. Luego de acudir a su médico de familia, éste le recetó tratamiento tópico con una serie de pomadas antibacterianas y anti-fúngicas. Sin embargo la úlcera no cicatrizó, razón por la cual fue remitido a un hospital para su estudio. En un primer momento los médicos pensaron en una posible neoplasia de piel. El estudio histopatológico de la biopsia no mostró sin embargo células neoplásicas pero si un tejido con células inflamatorias. Se repitió el análisis de la biopsia, pero esta vez en un centro especializado en neoplasias. El segundo reporte médico confirmó la ausencia de células malignas. Se decidió enviar la muestra a uno de los mejores centros europeos especializado en patología dermatológica. Allí, uno de los patólogos percibió unos pocos corpúsculos en uno que otro macrófago del paciente. Esto le trajo a la memoria lo que observó alguna vez en el curso internacional anual Gorgas del Instituto de Medicina Tropical "Alexander von Humboldt" del Perú. En aquél entonces se trataba de una biopsia de un paciente con Leishmaniasis Tegumentaria Americana. Con esa sospecha, sugirió que volviesen a interrogar al paciente sobre si estuvo alguna vez en alguna área endémica de Leishmaniasis. Efectivamente, el paciente estuvo de vacaciones en la selva de Cusco y Madre de Dios 15 ańos de atrás, en lo que hoy en día es la reserva nacional del Manu. El diagnóstico confirmatorio y la identificación de la especie de Leishmania fue realizada por técnicas de biología molecular, entre ellas la Polymerase Chain Reaction o PCR. El paciente fue subsecuentemente tratado con antimoniales y la úlcera desapareció al poco tiempo, aunque quedó preocupado pues fue informado que en algunos casos los pacientes de Leishmaniasis presentan metástasis en las mucosas oro-naso-faríngeas. La confirmación de la cura parasitológica se realizó posteriormente por la técnica de PCR, lo que le devolvió la tranquilidad En este ejemplo se nota claramente la influencia de la experiencia médica en el sesgo que inicialmente pueda tener el diagnóstico. En un país europeo, ante la ausencia de un cuestionario detallado, una úlcera de origen tropical no sería la primera opción para el diagnóstico. Fue la experiencia de un patólogo que observó en alguna oportunidad una imagen de parásitos intracelulares lo que inició la pista para el diagnóstico correcto. Igual de importante fue el aporte del laboratorio que incorporó el uso de técnicas de biología molecular; aquello que permitió conocer con precisión la causa de la lesión y evaluar posteriormente la cura post terapéutica. En esta última etapa, es la detección molecular la que se requiere para saber si el tratamiento logró eliminar los parásitos Leishmania porque los estudios de microscopía están por debajo de la sensibilidad requerida para tal fin. El diagnóstico de laboratorio moderno no sólo ofrece la ventaja de definir de manera objetiva el origen de la enfermedad, sino que además abre la posibilidad de enriquecer la información disponible. En el caso arriba descrito, se pudo identificar fehacientemente al microorganismo causante de la lesión. Adicionalmente, la caracterización molecular del ADN permite nutrir con datos específicos los estudios epidemiológicos de los países en que esa enfermedad es endémica. En el laboratorio moderno, las técnicas de biología molecular son usadas cada vez con mayor frecuencia. Ellas permiten no sólo caracterizar mejor al agente causal de la enfermedad, sino también el comprender mejor los mecanismos patológicos a nivel molecular. Debido a que las técnicas de biología molecular son una herramienta fundamental en la práctica de la medicina moderna, el presente artículo describirá los fundamentos y las estrategias del diagnóstico de enfermedades infecciosas basado en ellas. También se presentará algunos de los kits de diagnóstico de enfermedades infecciosas que son de utilidad en la práctica médica. Moléculas blanco para la detección del origen de la enfermedad El observar directamente al agente etiológico de una enfermedad es prueba irrefutable de la causa de la misma. Sin embargo, esto no siempre es posible, tal como ocurre en el caso de los virus o de la bacteria Mycobacterium leprae, causante de la Enfermedad de Hansen. Otras veces el tiempo de multiplicación celular en un cultivo es excesivo respecto a las necesidades prácticas para dar inicio a un tratamiento (ej: Mycobacterium tuberculosis). En otras ocasiones, el instrumental necesario para detectar el patógeno escapa a cualquier posibilidad de un centro de salud (ej: identificación de virus por microscopía electrónica luego de infección en cultivo de células eucariotes). Frente a la imposibilidad de observar u obtener un microorganismo completo, la tecnología moderna ha recurrido a detectar algún componente propio y exclusivo del patógeno. El componente que permite la identificación no debe estar presente en el humano hospedero ni tampoco debe ser compartido con otros microorganismos no patógenos. Si bien son los polipéptidos y los ácidos nucleícos aquellas macromoléculas para los cuales se han desarrollado procedimientos de detección, es el ADN el que ofrece inmejorables ventajas como blanco para el diagnóstico. En primer lugar, es la molécula presente en todos los seres vivos, excepto los virus ARN. Segundo, es químicamente estable, lo que permite su manipulación y almacenamiento. Tercero, es la base molecular de la identidad, lo que permite el diagnóstico certero. Cuarto, es el origen de la diversidad, por lo que se puede emplear para estudiar la contribución de la diversidad genética de patógenos de una especie a las variedades de formas clínicas de una enfermedad. żPor qué
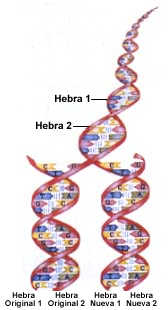
se usan preferentemente ácidos nucleicos para el diagnóstico molecular? El principio sobre el cual se basa el uso de ácidos nucleicos para el diagnóstico molecular es la complementaridad de secuencias entre hebras independientes que forman espontáneamente estructuras de doble hélice (Ver Figura 1). Esto ocurre por la interacción no covalente de pares Adenina- Timina (o con Uracilo en el ARN) y Guanina-Citosina. Cada par de estos nucleótidos por separado es incapaz de formar interacciones estables a las temperaturas de los seres vivos. Los nucleótidos, sin embargo, se encuentran formando parte de polímeros largos de ácidos nucleícos de tal manera que dos secuencias que sean complementarias se unirán mediante la sumatoria de interacciones débiles. El mejor ejemplo por analogía es el cierre de una maleta, donde cada diente de la cremallera es una pieza frágil pero que el conjunto de ellos contribuye a unir muy fuertemente los bordes de la maleta. En el caso del ADN, se requiere de temperaturas elevadas para separar las hebras complementarias (desnaturalización del ADN). Mientras más largo sean las porciones que formen enlaces débiles se obtendrá una mayor estabilidad. Si se disminuye paulatinamente la temperatura de la solución con el ADN desnaturalizado, las hebras separadas volverán a juntarse para reconstituir la doble hélice original. La complementaridad de dos hebras es la base del uso de sondas de ácidos nucleicos. Estas no son otra cosa que polímeros de nucleótidos con una secuencia específica que tienen una marca que permite su detección. Esta marca puede ser la incorporación en la molécula de un análogo químico radioactivo o fluorescente, o la unión covalente de una enzima que cataliza una reacción colorimétrica o quimioluminescente. Esta marca permite evidenciar la presencia de un híbrido entre una secuencia blanco y la sonda que interactúa con ella (Figura 2).
La formación de una doble hélice no requiere que las secuencias de las hebras que interactúan sean perfectamente complementarias. Ellas pueden presentar una proporción considerable de malos apareamientos ("mismatches") y a pesar de ello se formarán regiones de doble hélice siempre y cuando la estabilidad térmica se lo permita. Mientras más larga sea una secuencia, existirá una mayor posibilidad de formar malos apareamientos con secuencias que no son estrictamente complementarias. Esto explica el porque es conveniente utilizar secuencias de ADN relativamente cortas para usarlas como herramienta de diagnóstico y evitar de esta manera reacciones inespecíficas que resulten en diagnósticos falsos positivos. żQué secuencias
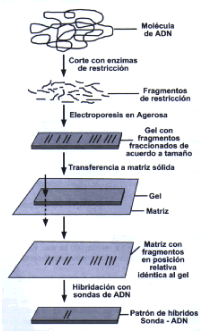
se utilizan como blanco para la identificación del agente patógeno? Existen por otro lado secuencias que no son compartidas por todos los individuos de una especie sino que se relacionan a sub grupos dentro de esa misma especie. Esta variabilidad del genoma se origina en las mutaciones puntuales o en los re-arreglos cromosómicos. Esta propiedad es inherente a la estructura de los ácidos nucleicos. Algunas de las secuencias están asociadas a características de patogenicidad y virulencia. La identificación de éstas secuencias es entonces también de interés médico. Aquí también no toda variante genética es de utilidad, por lo que se requiere de establecer de manera incontestable que ese marcador genético presenta una asociación directa. Métodos de detección de ADN El diagnóstico basado en el uso del ADN se realiza a través de la detección de secuencias de nucleótidos específicas que son propias para cada especie del patógeno. Históricamente la detección de secuencias se inició mediante técnicas de hibridación. Posteriormente, esta estrategia fue reemplazada por procedimientos que amplifican la secuencia blanco, la más conocida hoy en día, la PCR, siglas que provienen de la frase en inglés Polymerase Chain Reaction, y cuyos fundamentos veremos más adelante. Siguiendo el orden cronológico pasaremos a describir brevemente las tecnologías empleadas por laboratorios dedicados al diagnóstico molecular, así como el mencionar sus ventajas y desventajas. Diagnóstico por Southern blot e hibridación con sondas específicas. La hibridación consiste en la unión de una sonda de ADN con secuencias complementarias (Figura 3). Usualmente en este procedimiento, la sonda marcada se encuentra en solución y la molécula blanco casi siempre estará fijada a una matriz sólida. Hoy en día la de mayor uso es la matriz de Nylon, usualmente cargada positivamente, lo que contribuye a la unión de las moléculas de ADN cargadas negativamente. En esa matriz se coloca directamente la muestra a analizar donde se supone esta presente el patógeno, o se transfiere el ADN que ha sido previamente cortado con enzimas de restricción y luego separados por una electroforesis de acuerdo a su tamańo. Esta última técnica se conoce como la técnica de Southern blot, la que además de detección por hibridación, incrementa la especificidad por proporcionar un segundo elemento de juicio para el diagnóstico, que es la presencia de un fragmento de tamańo definido. El ADN blanco fijado sobre la superficie de una membrana de Nylon es tratado de tal forma que la interacción con la sonda es sólo posible a través de las secuencias de ADN expuestas. Esto determina que la sonda sólo puede unirse a la membrana de Nylon si y sólo si encuentra una hebra complementaria en su superficie. Mediante lavados de la membrana de Nylon con soluciones acuosas calientes se remueve toda sonda de ADN que no está unida a la secuencia de blanco. A mayor temperatura de lavado, mayor especificidad de detección. La detección del híbrido formado es indirecto. Si la sonda es radioactiva, entonces el híbrido se detecta por la impresión de un film sensible a la radiación emitida por el decaimiento del radioisótopo. Si en cambio se usa una sustancia cromógena en la sonda, la hibridación de ésta se hace evidente mediante una reacción enzimática ligada a la sonda. El producto de la reacción convierte al sustrato en un precipitado coloreado. Una variación de esta última estrategia es el uso de sustratos enzimáticos quimioluminiscentes los que al generar luz imprimen películas sensibles a la luz. Finalmente se revela la película impresa ya sea por radiación o quimioluminescencia o se desarrolla color en el mismo nylon que contiene la sonda cromogénica. Es obvio que la selección de la sonda adecuada para el diagnóstico es de crucial importancia, sin embargo, el desarrollo de las sondas específicas no será tratado en esta breve revisión a pesar que ello sea la parte más difícil y costosa. Si bien la especificidad es un enorme atributo de las sondas, su principal desventaja es que requieren de una cantidad elevada de ADN (aproximadamente un millón de copias), lo que corresponde a un número de microorganismos que sobrepasa lo que normalmente se encuentra en la muestra de pacientes. Además, se requiere de un tiempo relativamente prolongado, aunque menor que los cultivos, y de personal debidamente entrenado (1). Hoy en día, las técnicas de hibridación son rara vez usadas para el diagnóstico de manera independiente. Por lo general se usa en combinación con otras técnicas de detección. Las sondas son en cambio empleadas para la identificación de patógenos. Destacan las sondas quimioluminescentes que detectan secuencias específicas de ARN ribosomal que permiten identificar Mycobacterium tuberculosis, Mycobacterium avium (1), Histoplasma capsulatum y Cryptococcus neoformans (2,3). En aquellos casos que se usaron para el diagnóstico en muestras obtenidas de pacientes en el caso de infecciones por Chlamydia trachomatis y Neisseria gonorrhoeae (4,5), los niveles de sensibilidad y especificidad para la identificación fueron muy altos: 93.9% y 99.1% respectivamente para el caso de Chlamydia y de 93.3% y 99.0% para el caso de gonorrea. Diagnóstico por amplificación enzimática o PCR Como alternativa a la escasez de patógenos en presentes en la muestra del paciente, se planteó el incrementar el número de secuencias blancos, es decir, amplificar selectivamente una secuencia de todo el genoma en un tubo de ensayo.
Fue en 1987 que
se describe el procedimiento de Polymerase Chain Reaction o PCR con la
enzima de la bacteria termófila Thermus aquaticus. Las enzimas de esta
bacteria que vive a altas temperaturas, son muy termoestables. Es así
que su ADN polimerasa, la enzima responsable de la duplicación del ADN,
puede sintetizar la incorporación de nucleótidos a elevadas temperaturas,
teniendo un óptimo de 72° C y resistiendo la desnaturalización hasta 95°
C.
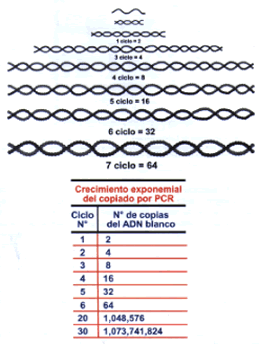
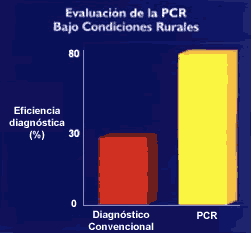
A continuación un calentamiento de la solución abrirá las hebras del ADN, las que al ser enfriadas nuevamente permitirán el anclaje de los cebadores que se encuentran en exceso. El anclaje de los cebadores también ocurrirá sobre las hebras recién sintetizadas y la Taq polimerasa volverá a copiar, tanto las hebras originales como las copias. Debido a la naturaleza del proceso de replicación que requiere de un molde, las cadenas sintetizadas tendrán un tamańo definido, correspondiente al tamańo del fragmento flanqueado por los cebadores. La duplicación de la secuencia blanco ocurre de una manera exponencial (Figura 5), es decir, primero 2 copias, luego 4, en la siguiente 16 y más tarde 32. Al cabo de 30 repeticiones, conocidos como ciclos, se habrá producido la amplificación masiva hasta bordear el millón de copias. Este número es ya visible por técnicas que usan colorantes afines a la doble hélice del ADN o de la hibridación con sondas en sistemas similares a los arriba descritos. Por lo tanto, una vez identificada y seleccionada una secuencia de ADN blanco, esta puede ser considerada para el desarrollo de un kit diagnóstico. El primer paso es determinar la sensibilidad y especificidad de detección (Figura 6). Es decir, cual es el número más bajo de patógenos detectados y si existe alguna otra secuencia de otros microorganismos que no siendo el patógeno buscado reaccionen con el PCR como si lo fueran. El principal problema de la PCR es la contaminación con ADN exógeno. Esto se debe a que esta tecnología es capaz de detectar muy pocas moléculas (5 o más copias), por lo que en un ambiente donde abunde el material patógeno o donde se realice intensamente actividades de detección por PCR, como es el caso de laboratorio de diagnóstico, se producen situaciones (principalmente de aerosoles) que favorecen la contaminación. Este problema conduce a resultados falsos positivos. Se evita este problema mediante la selección de personal altamente capacitado, el uso de ambientes específicos y restringidos y la adición de algunos productos químicos que convierten a los productos de amplificación en productos lábiles. El segundo paso a ser desarrollado es el determinar la sensibilidad diagnóstica, la especificidad diagnóstica, el valor predictivo positivo y el valor predictivo negativo. Para ello se requiere de pacientes comprobadamente diagnósticados por otros métodos directos (cultivo u observación directo) y otro conjunto de pacientes con signos y síntomas similares pero cuya enfermedad haya sido debido a otra causa comprobada por otros métodos de diagnóstico. Sólo una baja proporción de cebadores que han mostrado éxitos en el laboratorio tienen éxito en la práctica médica. Nuestra experiencia en el diagnóstico de la Leishmaniasis mediante la PCR nos muestra claramente que este procedimiento es superior a los métodos convencionales de cultivo y/o frotis (8). Incluso lo hemos llevado a condiciones de establecimientos de salud en áreas rurales (Figura 7). La PCR no requiere de las condiciones asépticas que requieren los cultivos, lo que representa una enorme ventaja para el diagnóstico de ésta enfermedad. Kits de diagnóstico basados en amplificación molecular in vitro aprobados por el Food & Drug Administration (FDA) de los Estados Unidos y de uso comercial. El panel de kits disponibles que han sido aprobados por el FDA es la resultante de rigurosos ensayos de validación. Este grupo de kits es una muestra representativa a nivel mundial de lo que el médico dispone, siempre y cuando sea realizado en laboratorios que reúnan ciertos requisitos. En la tabla 1 se muestran los kits validados hasta en ańo 1999 inclusive y otros que están próximos a a ser aceptados como herramienta de diagnóstico (mostrados con *). En la tercera columna de la Tabla 1 se indican los laboratorios que han desarrollado los sistemas de diagnóstico. Existen otros métodos de amplificación, como son LCR, TAS/3SR y QB replicasa, los cuales están en diferentes grados de desarrollo. El caso de LCR es el más avanzado, sobre todo en su aplicación a C. trachomatis y N. gonorrhoeae (9, 10), sin embargo, el elevado costo del equipo ha restringido su uso. En la tabla arriba mencionada, este procedimiento es mencionado como LCX.
Un campo particularmente notable de aplicación de PCR como herramienta de toma de decisiones es el de carga viral. Es decir, no basta saber que un individuo esta infectado con el virus HIV sino establecer cual es la variación de la concentración de partículas virales en caso de ser sometidos a tratamiento. Actualmente las empresas Roche, Chiron y Organon disponen de métodos de cuantificación viral. En todos ellos se requiere de las condiciones óptimas que requiere todo laboratorio dedicado a detección molecular. Conclusiones Ha transcurrido un tiempo largo desde los primeros métodos de detección usando fragmentos de ADN como sondas. Los métodos de hibridación son empleados hoy con propósitos de identificación o de incrementar la sensibilidad de los kits de diagnóstico. Es la PCR, y otros métodos de amplificación, los que predominan en el panel de herramientas de diagnóstico. Sus elevados costos y la necesidad de personal muy bien entrenado, restringen su uso a enfermedades virales o aquellas situaciones en las que los métodos convencionales no permiten detectar al agente causal en un tiempo útil para la toma de decisiones médicas. Los métodos de amplificación molecular son los que se emplean hoy en día en los laboratorios de diagnóstico más modernos. La sensibilidad y especificidad compensan los costos relativamente elevados, sobre todo en aquellas enfermedades donde el cultivo o la serología son poco prácticos o imprecisos, respectivamente. Existen suficientes argumentos para pensar que en un próximo futuro los métodos de detección molecular desplazaran a los métodos convencionales. Lo que si es seguro, es que todo método a desarrollarse se basará en las propiedades químicas de la doble hélice del ADN. Bibliografía 1. López, M; Montoya, Y; Arana, M; Cruzalegui, F; Braga, J; Llanos-Cuentas, A; Romero, G. and Arévalo, J. The use of nonradioactive DNA probes for the characterizacion of Leishmania isolates from Peru. American Journal of Tropical Medicine and Hygiene 1988; 38: 308-314. 2. Goto, MS; Oka, S; Okuzumi, K. et al. Evaluation of acridinium-ester labeled DNA probes for identification of Mycobacterium tuberculosis and Mycobacterium avium-Mycobacterium intracellulare complex in culture. J Clin Microbiol 1991; 29: 2473-2476. 3. Huffnagle, KE; Gander, RM. Evaluation of Gen-Probe's Histoplasma capsulatum and Cryptococcus neoformans AccuProbes. J Clin Microbiol 1993; 31: 419-4219. 4. Padhye, AA; Smith, G; McLaughlin, D. et al. Comparative evaluation of a chemiluminescent DNA probe and an exoantigen test for rapid identification of Histoplasma capsulatum. J Clin Microbiol 1992; 30:3108-3111. 5. Hosein, IK; Kaunitz, AM; Craft, SJ. Detection of cervical Chlamydia trachomatis and Neisseria gonorrhoeae with deoxyribonucleic acid probe assays in obstetric patients. Am J Obstet Gynecol 1992;167:588-591. 6. Limberger, RJ; Biega, R; Evancoe, A. et al. Evaluation of culture and the Gen-Probe PACE 2 assay for detection of Neisseria gonorrhoeae and Chlamydia trachomatis in endocervical specimens transported to a state health laboratory. J Clin Microbiol 1992; 30:1162 -1166. 7. Mullis, KB. y Faloona, F. Specific síntesis of DNA in vitro via polymerase-catalyzed chain reaction. Methods Enzymol. 1987;155: 335-350. 8. Saiki, RK; Gelfand, DH; Stoffel, S; Scharf, S; Higuchi, R; Horn, GT; Mullis, KB. y Erlich, HA. Primer directed enzymatic amplification of DNA with a thermostable DNA polymerase. Science 1988; 239: 487- 491. 9. López, M; Orrego, C; Cangalaya, M; Inga, R; Arévalo, J. Diagnosis of Leishmania via the Polymerase Chain Reaction a Simplified Procedure for Field Work. Am. J. Trop. Med. Hyg. 1939; 4(3): 348-356. 10. Birkenmeyer, L. and Armstrong, AS. Preliminary evaluation of the ligase chain reaction for specific detection of Neisseria gonorrhoeae. J Clin Microbiol 1992; 30:3089-3094. 11. Dille, BJ; Butzen, CC. and Birkenmeyer, LG. Amplification of Chlamydia trachomatis DNA by ligase chain reaction. J Clin Microbiol 1993; 31:729-731. .
(*) Profesor Principal Dpto. de Ciencias Fisiológicas, Universidad Peruana Cayetano Heredia |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||